在比赛开始时,我们的身体必须从静息状态迅速过渡到高强度状态。
在比赛过程中,甚至在日常训练中,我们都需要通过改变强度来加速。
这些情况都要求机体快速响应代谢需求。我们的生理系统必须适应节奏的变化。
根据 Westerblad et al. (2002) 的研究,"骨骼肌细胞的能量消耗在从静息状态到高强度运动时可以增加到100 倍"。
在本文中,我们将解释氧气消耗(VO 2)动力学的概念,以及它在你应对运动强度快速变化能力中所起的作用。
能量供应系统回顾
ATP 是肌肉唯一能够直接使用的能量形式。然而,肌肉内储存的 ATP 只能维持几秒钟的运动,但幸运的是,ATP 可以通过多种途径重新合成,从而让运动得以持续。
理想情况下,肌肉能量需求的增加(从静息状态到运动状态)应该由有氧代谢系统的能量供应即时增加来满足(在有氧代谢中,氧气参与碳水化合物或脂肪氧化产生 ATP)。
然而,有氧代谢提供的能量(通过 VO 2 测量反映)增加得相对缓慢,根据训练水平的不同,大约需要 1 到 3 分钟才能达到稳定状态(Poole and Jones, 2012)。这可能是由于肌肉内部过程的滞后性,以及有氧系统产生 ATP 所需的步骤比其他系统更多。
氧气消耗动力学与氧亏
运动开始后氧气消耗的变化过程被称为"VO 2 动力学",VO 2 动力学的快慢由 VO 2 达到稳定状态的速度来定义。
由于 VO 2 动力学相对缓慢,至少相对于肌肉能量需求的即时增加而言,必须启动其他能量产生途径来满足需求。
这种需求的大小可以用"氧亏"(oxygen deficit,O 2 亏损)的概念来说明。氧亏简单来说就是在一定时间内以目标强度进行运动所需的能量总量,与同一时期内有氧代谢实际提供的能量量之间的差值。
因此我们可以推断,相当于氧亏的这部分能量必须几乎完全由无氧代谢过程来提供(启用磷酸原系统和糖酵解系统)。
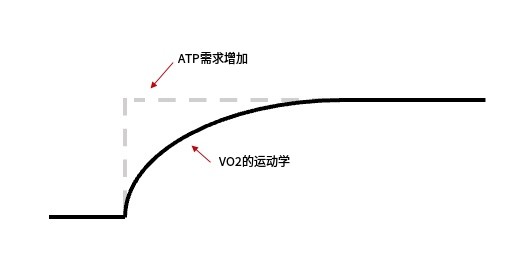
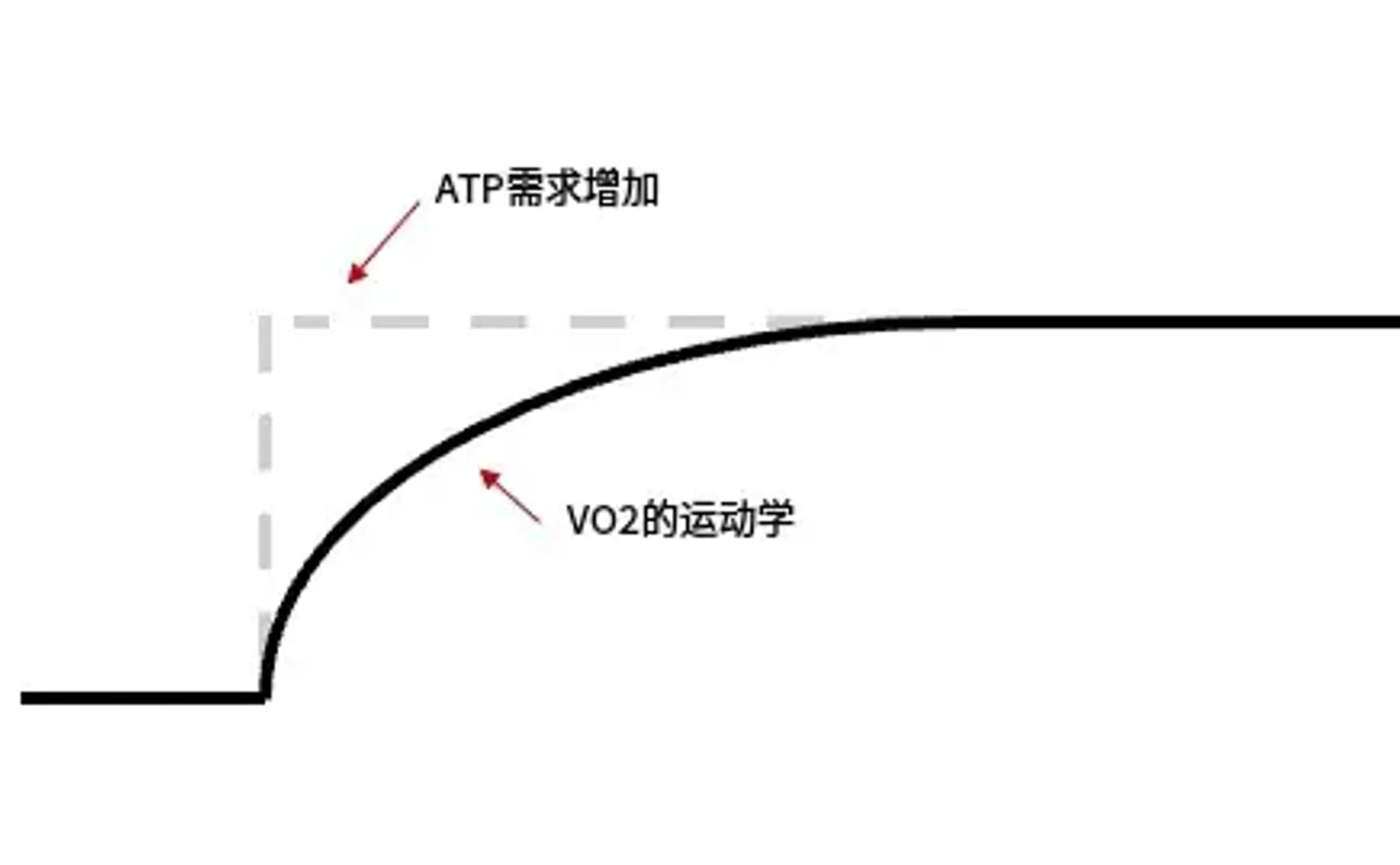
图 1:恒定强度运动开始后 VO 2 响应示意图。收缩肌肉细胞中的 ATP 需求急剧上升,但 VO 2 的响应(反映有氧代谢提供的能量)增加相对缓慢,需要几分钟才能达到所需的平衡水平。虚线和实线之间的区域代表氧亏,对应于必须由无氧代谢过程提供的能量量。
氧亏过大的后果
人体还有另一种快速可用的能量储备——磷酸肌酸(PCr)。然而,和 ATP 一样,这种资源也非常有限。因此,机体不得不增加对糖酵解系统的依赖来满足部分能量需求。
尽管这些无氧代谢机制(磷酸原系统和糖酵解系统)对于在有氧代谢率持续上升至所需水平期间维持运动至关重要,但它们的使用会带来负面后果。
氧亏越大,高能磷酸化合物(ATP、PCr)的分解就越多,糖酵解系统的激活程度就越高,这会导致收缩肌肉中 PCr 和 ATP 浓度下降,以及代谢副产物如 ADP、无机磷酸盐、氢离子浓度的增加。此外,由于糖酵解系统是一个相对低效的过程,肌糖原储备的消耗速度会比通过有氧氧化产生相同量 ATP 时快得多。
所有这些因素都与神经肌肉疲劳过程密切相关,因此,在运动最初几分钟内积累大量氧亏会对耐力表现产生负面影响。即使在马拉松这样的项目中,产生初始氧亏的 1 到 3 分钟可能只占总运动时间的 1%到 2%,但代谢产物的积累会负面改变肌肉细胞内建立的生化环境,从而影响比赛后期的表现(MacIntosh, Brian R et al., 2012)。
研究表明,VO 2 动力学快的运动员能够更早达到 VO 2 max,并且积累的氧亏较小(Niemeyer, Max et al., 2019)。一旦达到 VO 2 max,他们能够继续维持高强度运动,能够在 VO 2 max 水平上稳定持续。相反,VO 2 动力学慢的运动员会积累大量氧亏。一旦达到 VO 2 max,他们就无法继续维持运动,因为他们已经积累了大量代谢产物,并耗尽了高能磷酸化合物储备(ATP、PCr)。
最后,研究报告显示 VO 2 动力学的时间常数与第二阈值显著相关(Murgatroyd, Scott R et al., 2011),这表明这两个变量可能通过共同的生理机制相关联,如骼肌的有氧氧化能力(Whipp, B J., 1987)、高度氧化的 I 型纤维比例(Vanhatalo, Anni et al., 2016)以及毛细血管密度(Mitchell, Emma A et al., 2018)。
VO 2 动力学在运动表现中的作用
现在很明显,有氧代谢系统活性提升得越快(即 VO 2 动力学越快),运动表现就应该越好。
事实上,精英耐力运动员拥有极快的 VO 2 动力学,而缺乏运动的人群、老年人和患病人群的 VO 2 动力学则慢得多,这绝非偶然(Rossiter, 2011)。
耐克 Breaking 2 项目(旨在让人类马拉松突破 2 小时大关)的研究人员报告了 VO 2 动力学时间常数与马拉松成绩之间的正相关关系(Jones, Andrew M et al., 2021)。换句话说,VO 2 动力学越快,马拉松成绩就越好。
因此,VO 2 动力学是耐力运动的重要表现因素。它也很可能是团队运动或间歇性运动的表现因素,因为这些运动中代谢需求的变化非常剧烈。
事实上,快速的动力学还能让运动员在多次努力之间更好地恢复。积累氧亏最少的运动员能够更快地恢复到基线水平,因为他们没有需要偿还的氧债(oxygen debt)。
东非长跑运动员通常具有非常快的 VO 2 动力学,同时伴随着出色的跑步经济性。这很可能就是为什么他们的 VO 2 max 并不是特别高(与同等水平的欧洲选手相比)的原因(Jones, Andrew M et al., 2021)。
最后,改善 VO 2 动力学对训练也可能很重要。例如,旨在提高 VO 2 max 的训练课通常目标是累积尽可能多的接近 VO 2 max 的训练时间。如果有氧系统能够更快地做出响应,那么在给定的训练负荷下,就能在 VO 2 max 附近累积更多的时间,从而使训练课更加高效。
时间常数
VO 2 响应在运动开始后的最初几分钟内遵循近似指数函数的时间变化规律,而指数响应可以用所谓的"时间常数"(τ,tau,表示达到稳定状态所需时间的指标)进行数学表征。
指数过程通常用数学方式定义,当它们在图表上随时间绘制时,呈现出特征性的增长或衰减曲线。不过,你可以简单地将它们理解为:在任何给定时刻,增长或衰减的速率与该时刻的现有量相关。
当经过四个时间常数后,指数过程被认为基本完成。因此, 对于 VO 2 动力学来说,如果时间常数为 40 秒(这是健康但未经训练人群的典型值),意味着 VO 2 的"稳定状态"将在大约 160 秒内达到。相比之下,据报道精英耐力运动员的时间常数为 15 秒或更短(稳定状态在约 40 到 60 秒内达到),而患有晚期心血管、肺部或代谢疾病的患者时间常数可达 90 秒(稳定状态在约 6 分钟内达到)(Poole, David C, and Andrew M Jones., 2012)!
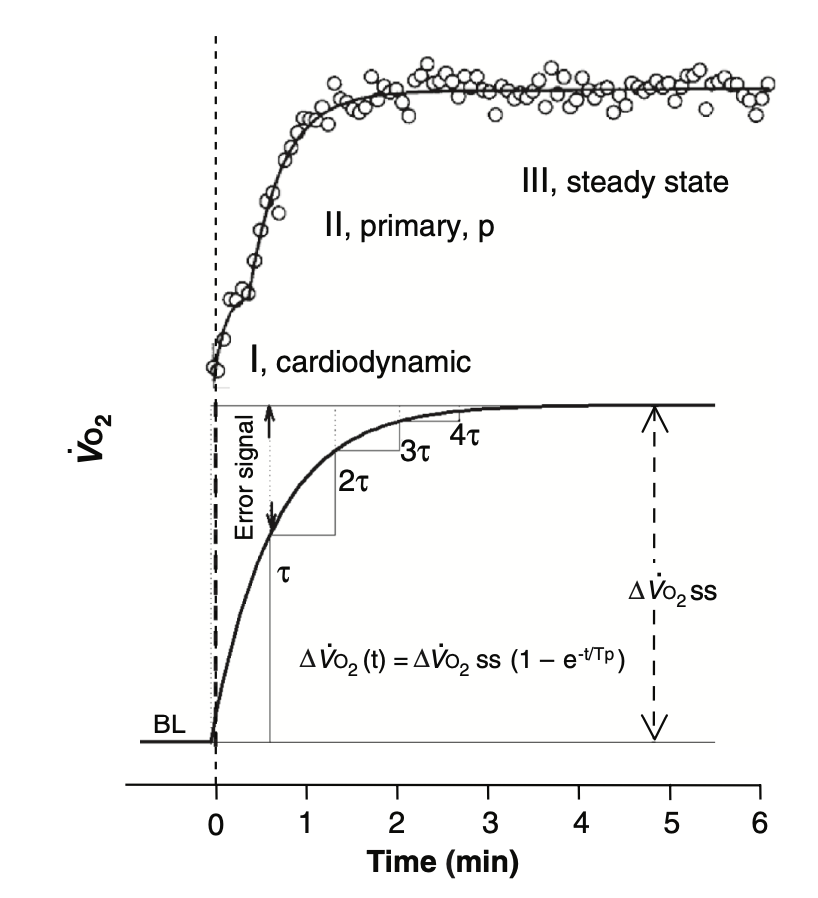
对于相同的代谢率增幅(从静息水平开始),高水平运动员产生的氧亏仅为年轻久坐人群的四分之一到三分之一,仅为患者的八分之一到六分之一。
尽管其他表现因素如 VO 2 max、运动经济性和乳酸阈值也能区分这三类人群,但 VO 2 动力学对于解释运动表现差异的重要性不应被低估。事实上,对于患者来说,极慢的 VO 2 动力学会导致即使在轻度运动时也会快速出现肌肉疲劳,从而妨碍他们舒适地完成日常生活活动。对于耐力运动员来说,更快地达到比赛所需的 VO 2 不仅能减少可能损害表现的代谢产物积累,还能"节省"一部分有限的高能磷酸化合物(ATP、PCr)储备,以便在比赛后期使用,比如在终点冲刺时。
结论
氧气消耗动力学的快慢是一个重要但容易被忽视的表现因素。
一般来说,VO 2 动力学对于持续 2 到 40 分钟的项目,或者从起跑线开始就需要超阈值强度的项目尤为重要。如果你参加这类项目,就应该关注你的 VO 2 动力学!
参考文献
-
Jones, A. M., Kirby, B. S., Clark, I. E., Rice, H. M., Fulkerson, E., Wylie, L. J., Wilkerson, D. P., Vanhatalo, A., & Wilkins, B. W. (2021). Physiological demands of running at 2-hour marathon race pace. Journal of Applied Physiology, 130(2), 369-379. https://doi.org/10.1152/japplphysiol.00647.2020
-
MacIntosh, B. R., Holash, R. J., & Renaud, J. M. (2012). Skeletal muscle fatigue–regulation of excitation-contraction coupling to avoid metabolic catastrophe. Journal of Cell Science, 125(Pt 9), 2105-2114. https://doi.org/10.1242/jcs.093674
-
Mitchell, E. A., Martin, N. R. W., Turner, M. C., Taylor, C. W., & Gropper, H. (2018). Critical power is positively related to skeletal muscle capillarity and type I muscle fibers in endurance-trained individuals. Journal of Applied Physiology, 125(3), 737-745. https://doi.org/10.1152/japplphysiol.01126.2017
-
Murgatroyd, S. R., Ferguson, C., Ward, S. A., Whipp, B. J., & Rossiter, H. B. (2011). Pulmonary O₂ uptake kinetics as a determinant of high-intensity exercise tolerance in humans. Journal of Applied Physiology, 110(6), 1598-1606. https://doi.org/10.1152/japplphysiol.01092.2010
-
Niemeyer, M., Leithäuser, R. M., Beneke, R., & Schümacher, Y. O. (2019). Oxygen uptake plateau occurrence depends on oxygen kinetics and oxygen deficit accumulation. Scandinavian Journal of Medicine & Science in Sports, 29(10), 1466-1472. https://doi.org/10.1111/sms.13493
-
Poole, D. C., & Jones, A. M. (2012). Oxygen uptake kinetics. Comprehensive Physiology, 2(2), 933-996. https://doi.org/10.1002/cphy.c100072
-
Rossiter, H. B. (2011). Exercise: Kinetic considerations for gas exchange. Comprehensive Physiology, 1(1), 203-244. https://doi.org/10.1002/cphy.c090010
-
Vanhatalo, A., Fulford, J., DiMenna, F. J., & Jones, A. M. (2016). The mechanistic bases of the power-time relationship: muscle metabolic responses and relationships to muscle fibre type. The Journal of Physiology, 594(15), 4407-4423. https://doi.org/10.1113/JP271879
-
Westerblad, H., Allen, D. G., & Lännergren, J. (2002). Muscle fatigue: lactic acid or inorganic phosphate the major cause? News in Physiological Sciences, 17, 17-21. https://doi.org/10.1152/physiologyonline.2002.17.1.17
-
Whipp, B. J. (1987). Dynamics of pulmonary gas exchange. Circulation, 76(6 Pt 2), VI 18-VI 28.